Life Sciences Centre
2350 Health Sciences Mall
Vancouver, BC V6T 1Z3
A MESSAGE FROM THE DEPARTMENT HEAD by Mike Gold
In the great cycle of (academic) life, our Department is always welcoming new students and faculty while others become part of our alumni family. This year we had two of our professors, George Spiegelman and Hung-Sia Teh ‘graduate’ to retirement, each with plans that will keep them extremely busy. At the same time, we welcomed two new faculty members, Martin Hirst and Georgia Perona-Wright. Martin spent the previous few years at the BC Genome Sciences Centre and in this post-genomic era is bringing epigenetics to our department. Georgia is originally from the UK and worked at the Trudeau Institute in the woods of upstate New York before joining us. She is an ‘integrative immunologist’ who is interested in determining how local versus systemic immune responses determine whether or not different kinds of infections are cleared. She brought with her to Vancouver many interesting infectious disease models as well as her own canoe. We are currently in the process of hiring two more faculty members in exciting new, cross-disciplinary areas, geomicrobiology and human microbial ecology. The geomicrobiologist is a joint hire with the Department of Earth & Ocean Sciences. They will bring new dimensions to our environmental microbiology research and will further develop our combined undergraduate program in oceanography and microbiology. Microbial ecology originated as an aspect of environmental microbiology and it is now realized that the approaches developed to study microbial communities in the oceans or soils can be applied to the microbial communities of our gut, lung, skin, etc. These microbial communities and their interactions with the host are key determinants of health and disease. The combined efforts of microbiologists, immunologists, genome scientists, and bioinformaticians will be required to fully understand these complex interactions.
The fields of microbiology and immunology will look very different 20 years from now, just as the microbiology and immunology that I learned as a students seems like ancient history to our current students. That’s a good thing and we’re ready for it, not only with new faculty members but also with graduate students and undergraduates who teach us new things all the time. Our goal as faculty members is to train our students to learn on their own, explore the world of science, come up with their own ideas, and make new connections. The evolving nature of how we teach our undergraduate courses and how we train our graduate students seems to be achieving this goal. My job is to try to keep up with my students. There is so much to know and no one person can know it all. In our labs, our classes, and our lives we have to create human internets in which everyone’s knowledge is connected so that we can draw on the collective knowledge and activities of the community, just like the distributed metabolic pathways that allow microbial communities to generate the nutrients and metabolites that each individual species needs. To succeed in this, we all have to be open to new information and be active, life-long learners. Recently I’ve been reading a book called “An Incomplete Education” and it’s convinced me that my education is an on-going process that is still very much incomplete.
Every year I look forward to celebrating the accomplishments of our graduating students, both at graduation and at the graduation banquet. The challenge each year is to come up with a new, and hopefully inspiring speech, for the graduation banquet. Unfortunately I was not able to attend the banquet this year as I was in Paris. Quel dommage! However, Rachel Fernandez filled in ably for me. In thinking about what I might have said (I’m writing it down here so I can remember it for next year), several quotes have been going through my head recently about how one needs to be an active agent of change in your own life and in the world. The first two quotes are from Robert F. Kennedy Jr., who said: “There are those that look at things the way they are, and ask why? I dream of things that never were, and ask why not?” and “Only those who dare to fail greatly can ever achieve greatly.” The other quote I like a very much is from a radio news reporter named Scoop Nisker that I listened to when I lived in Berkeley, California. His sign-off line was “If you don’t like the news, go out and make some of your own”. It’s up to each of use to do that in our own way, and to make a difference to those around us. Quoting Robert F. Kennedy again, “The purpose of life is to contribute in some way to making things better.”
A TIME TO WORK AND A TIME TO PLAY by Julyet Benbasat
If you ever see me driving “no hands”, just get out of the way - I earned my license in Istanbul eons ago! These days, in a contemplative mood upon my retirement, I often drive down memory lane wondering how I got half way around the world to Vancouver:
University has been a huge part of my life both as a student and faculty member – 47 years to be exact! Waiting in line for registration to Chemical Engineering at Robert College in the heights above the Bosphorus in Istanbul is how I met my husband Izak. He said: “this is where I went to high school; let me show you around.” He did. :)
I loved the second year labs, but wearing the latest fashion dress my grandparents had just brought from Paris was not the best idea. A drop of whatever it was, and the dress essentially disappeared off of me. Easy decision: I won’t be an organic chemist! Lesson learned: always button up my lab coat!
Biochemistry was all the rage when I was admitted for a doctorate at the University of Minnesota. I studied the assembly of bacterial viruses from their parts. I selected mutants that allowed purification and concentration of individual parts like heads, tails, tail fibers, then derived the theory and algorithms for interpretation of the assembly kinetics observed by inelastic light scattering. The cold winters were perfect for concentrating on work. The fact that I was able to repeat all the experiments that went into the thesis in just two weeks before the defense gave me pause, but I didn’t dwell on it; I told myself this was normal!
PhD at hand, UBC was an easy choice: In a mid February interview, the weather was mild, snowdrops blooming, sun shining – I was humming while waiting at the bus stop! I accepted a postdoc appointment in Chemistry, studying inhibitors of acetylcholinesterase. We isolated the enzyme from 6 foot eels imported from the Amazon. One morning I walked in, saw the aquarium empty and the eel sprawled on the floor of the lab…That’s when I decided I would stick to teaching and to researching small things, like viruses.
After a 3 year stint in Biochemistry (BioC 403, Enzymology) I continued my career in the Microbiology and Immunology Department. Starting with the large Biol 201 and Micb 200 courses, I must have taught 1000 students a year for the first 12 years leading to a Killam Teaching Prize. One day, I complained to the Dean that things were getting too routine; glass ceiling and all. He called a month later: Would I want to be the director of the brand new Science One Program? Absolutely! Check a box: If you want change, say so! With that started six totally inspirational years in creating an interdisciplinary learning environment for first year students looking for a challenge, teaching with an amazing team of faculty, and negotiating everything from space to funding to faculty recruitment from departments. My son says this was an engineering tour de force…. right along the lines of my undergraduate education. The program certainly felt right and it wasn’t easy to leave it to a new director, but I was ready to be associate dean to apply what I had learned from the former experience to set up the Integrated Sciences and Coordinated Sciences Programs.
Over the last several years in Microbiology and Immunology, Micb 322, Biol 112 and Micb 202 occupied my thoughts and time. I am ready for “retirement” now and hope that I will be busier than ever still learning, travelling and still waking up looking forward to the coming day. What will I miss most? Being part of this community full of students and the energy they bring.
Ada Mew is the outgoing President of MISA, and incoming VP Finance. Rhena Yoo is the outgoing Communication and Marketing director, graduating in May 2013 with an honors in Biotechnology.
The Microbiology Immunology Student Association, otherwise known as MISA, carries 27 years of traditions and responsibilities. Each year, we uphold the decade-old goal: promote a fun, academic and social atmosphere for microbiology enthusiasts. While the scope and number of our activities have changed over the years, the basis remains. We cater to the students by hosting a handful of social events throughout the year, and provide guidance through their journey here at UBC.
Through the social aspect, we've tried to connect the members of our great faculty. Events like the 'Welcome Back BBQ' is the icebreaker to meet professors, graduate students, and undergraduates. We provide the hot dog and atmosphere, they bring the conversations. Other exciting activities like Hockey Nights at Abdul Ladha's Science center helps our students wind down, while cheering for our favorite boys in blue, green and white.
Left: Crystal Lee welcomes the incoming and returning MBIM students with a free hotdog Right: Kevin Tsai grills hot dogs in preparation to feed starving students
On the academic end of the spectrum, we've hosted a couple of information session nights. Graduate students and faculty members are invited to speak about their current and recent experiences. Students not only hear about the latest advances happening right in their own backyard, but glean insight on how they themselves can get started early. This year, we hosted a 'Career Night' geared towards graduating students about working in industry, academia, and continuing studies in graduate work. We hope for more events to help our graduating undergrads transition into their next steps in life, such as resume-building and career services.
MISA Presents Research Night and the turnout was a great success!
The MISA Executive team has put in countless hours putting together many of these events, and keeping the community in the know. While majority are now departing from the UBC Microbiology community, we have a new executive team ready to build on the foundations passed on from this year. Their mission statement is to forge new connections with MISA and the outside community, to promote the interest of Microbiology to everyone! We trust they will continue to uphold the excellence of UBC's Microbiology faculty.
Before we bid adieu, we'd like to thank all those who came and consistently support the activities of our club! We'd like to thank the faculty, the students and all of UBC for the greatest pleasure of our lives. As Jonas Salk once said, "I feel that the greatest reward for doing is the opportunity to do more."
THE MURPHY LAB: A HARDCORE METAL GROUP by Catherine Gaudin and Michael Murphy
When red blood cells break down intravascularly due to the end of their life cycle or to injury, the iron-containing heme groups of hemoglobin (Hb) released into the plasma could take part in destructive redox reactions. However, an abundant serum protein called haptoglobin binds plasma Hb with high affinity and safely accompanies it to a macrophage or monocyte, where the complex is recognized by CD163 and engulfed. This is only one example of the numerous systems in place to control the availability of heme and other forms of iron in the human body, which have as much to do with preventing wanton destruction due to uncontrolled iron-catalyzed redox reactions as they do with limiting the amount of iron available to invading microbes. With such effective protective systems, how could an iron-requiring bacterium gain a foothold and cause disease?
The answer is: by using these very systems against us. Due to normal red cell turnover, the Hb-haptoglobin levels in human plasma can be in the range of 2 μM, yet the opportunistic pathogen Staphylococcus aureus grows happily with 0.5 μMHb as the sole iron source.
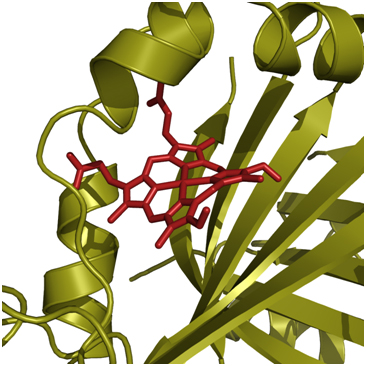
Under low iron conditions, S. aureus expresses surface proteins which are able to bind to Hb (IsdB and IsdH) as well as haptoglobin (IsdH), thereby acting as a sponge for any released Hb. IsdB and IsdH are part ofa system of heme and hemoprotein receptors known as the Iron-regulated surface determinant (Isd) system.The major Hb-binding component, IsdB, is able to remove the heme constituent from Hb and, through a series of reactions with other Isd proteins, transfer the heme to a membrane transporter where it can be internalized and used as an iron source by the bacterial cell. This fascinating and effective method of iron acquisition has been a focus of the Murphy Lab for several years.
Graduate students Jason Grigg and Catherine Gaudin characterized heme binding by two Isd proteins, IsdA and IsdB. They determined, using X-ray crystallography, heme transfer kinetics and site-directed mutagenesis, the mechanism by which a conserved domain present in varying copy number on the surface proteins, the NEAT domain, is able to bind and relay heme between surface Isd proteins.
More recently, the focus has been on the interaction between IsdB and host Hb. It is thought that a second IsdB NEAT domain is responsible for Hb binding; Catherine Gaudin and Meghan Verstraete (a talented directed studies student) have been working to either confirm or refute this. Armed with new static light scattering equipment and a plethora of IsdB constructs, Catherine and Meghan are probing which regions of IsdB interact with Hb, as well as how the different domains of IsdB work together to remove the heme molecules from Hb. Delving deep into the unfamiliar territory of Hb biochemistry has been much more complex than anticipated, but will eventually shape a model of this exciting aspect of host-pathogen interaction.
One further area of Isd system function that the Murphy Lab has played a major role in characterizing is the heme degradation pathway by two intracellular enzymes, the paralogs IsdG and IsdI. Work pioneered by Dr. Woo Cheol Lee (a past post-doc) and Georgia Ukpabi (a recent M. Sc. graduate) determined that IsdGI represent a totally new class of heme-degrading enzymes. In collaboration with the Skaar group at Vanderbilt University, we showed that IsdGI were completely different from the well-known heme oxygenases in protein fold, enzymatic mechanism and the heme breakdown product. The mechanism is not well understood, but the strange conformation of the heme in the binding pocket is believed to hold the key. The heme ring is highly contorted into a “ruffled” shape, like the potato chip (see inset). Our newest post-doc, Dr. Slade Loutet, will be continuing the work on how heme ruffling contributes to the enzyme activity. Since these proteins are so different from our own heme oxygenases, there is hope that someday they might be an effective therapeutic target in the fight against this insidious pathogen.
THE POWER IS IN THE NUMBERS...THE NUMBER OF ALLELES THAT IS. by Pamela Lincez and Marc Horwitz
With a simple change or single nucleotide polymorphism (SNP) in the sequence of a gene, variant alleles can form, fueling the power and beginning of a numbers game. Unless you’ve been hiding under a genetics rock, you are most likely aware that for almost a decade, genetic epidemiologists have been tracking the numbers of variant alleles across populations of individuals in genome-wide association studies (GWASs) in hopes of uncovering traits or risk factors associated with genetic variation.The number of variant alleles created when an individual is created, can decide the fate of that individual in the face of many of nature’s challenges including cancer, autoimmune disease, and aging. In the case of Type 1 Diabetes (T1D), an autoimmune disease prevalent in young children, a collection of risk alleles exist that can unfortunately dictate independently or as a collective, in conjunction with other non-genetic factors, the fate of disease onset. Out of the 50 or so risk alleles associated with T1D, theAla946 allele resulting from a common non-synonymous SNP (nsSNP) in the interferon induced with helicase C domain 1 (IFIH1) gene, a gene that expresses a protein important for viral sensing, has more recently been getting a lot of special attention. In 2006, a GWAS lead by John A. Todd in Cambridge, UK, identified a common coding variant in IFIH1 (rs1990760, A946T) of T1D patients, which provided statistical support for another T1D risk locus. Later, an allelic specific expression analysis study from the same group demonstrated the protective association of the Ala946 allele with T1D, correlated with less IFIH1 transcripts being expressed. This exciting and interesting discovery demonstrates not only the importance a threshold in a particular gene’s expression may have in conferring protection from autoimmune disease onset, but also implicates an anti-viral response gene in T1D. In our lab we model autoimmune diseases including T1D and Multiple Sclerosis in genetically susceptible mice using certain viruses to accelerate the onset of disease, so the discovery of the protective Ala946 allele in the T1D risk gene IFIH1 that expresses a virus sensing protein naturally became inspiration for some new projects.
T1D is a consequence from the destruction of pancreatic insulin-producing beta-cells. The autoimmune response that ensues in the pancreas leads to a significant depletion of insulin production, which in turn, causes uncontrollable and unused levels of glucose in the bloodstream. Though SNPs strongly influence an individual’s susceptibility to disease, they are not the sole contributors to the onset of disease-T1D is a multifactorial disease, where environmental factors such as diet and particular pathogen infections are as well strongly implicated. Enteroviruses, specifically the Coxsackie virus B (CVB) group, are the most likely T1D causative pathogens. CVBhas been isolated from the pancreas of acute onset patients, further adapted to mice and has shown to induce in mice, T1D-like pathologies seen in patients including hyperglycemia, the production of islet antigen autoantibodies, and islet beta-cell destruction. The demonstration of CVB’s potential to induce T1D pathologies in genetically susceptible mice gave rise to the widely accepted and central hypothesisin our labthat interplay between virus infection and host genetics induces the onset of autoimmune disease.
The T1D risk gene IFIH1 is responsible for expressing the protein commonly referred to as melanoma differentiation-associated gene-5 (MDA-5), which acts as an intracellular virus sensor in the cytoplasm of most cells, though most prevalently in immune cells. MDA-5 senses virus dsRNA and with CVB being a ssRNA virus that produces dsRNA as a replicative byproduct, it is likely that MDA-5 plays a critical role in CVB detection alongside other cytoplasmic dsRNA sensors such as retinoic acid-inducible gene I (RIG-I) and Toll-like receptor 3 (TLR3). Drawing inspiration from the discovery of the protective Ala946 allele in the MDA-5 gene IFIH1 and our lab’s previous work with CVB- induced T1D, our lab is interested in studying the immune effects of dampened MDA-5 expression, such as in the case with the protective allele, with CVB-induced T1D in the non-obese diabetic (NOD) mouse- a genetically susceptible mouse that spontaneously develops diabetes. In fact, we are working toward generating a transgenic NOD mouse that is mutated in the MDA-5 gene to carry the protective Ala946 allele. We anticipate that mice carrying the knock-in (KI)T946A mutation in the IFIH1 gene will be protected from autoimmune disease onset following CVB infection since the Ala946 allele will likely result in reduced IFIH1 gene expression and dampened anti-viral responses linked to the acceleration of disease. We are also working toward dampening MDA-5 expression and function using in vivo delivery of siRNA targeting MDA-5 and in vivo delivery of adenovirus-associated vectors carrying siRNA directed at MDA-5.
While working toward the generation of the NODMDA5T946A KI mice, we have successfully backcrossed MDA5 deficient mice onto the NOD background (NODMDA5ko), confirming their status with SNP analysis. We have also observed that NOD MDA5 heterozygotes (MDA-5 hets) and wild-type NOD (WT) siblings develop spontaneous autoimmune diabetes similarly. More importantly, NODMDA5ko mice do not develop spontaneous diabetes by 30 weeks of age and have impaired survival following high dose CVB challenge compared to MDA-5 hets and WT mice. This suggests that a lack of any functional MDA-5 severely impairs anti-CBV immune defenses. As patient polymorphisms in MDA-5 such as the common nsSNP A946T likely retain some level of MDA-5 function, we have continued our analysis of the immune response in CBV- induced T1D in MDA-5 hets. Preliminary studies examining inflammatory cytokines and adaptive immune responses following CBV infection as well as proliferation of diabetogenic T cells in MDA-5 het recipients have suggested a distinct phenotype for reduced MDA-5 expression compared to MDA-5 KOs and WT mice. Dampened MDA-5 expression seems to alter early inflammatory signaling and changes the type of adaptive response that can ultimately lead to T1D protection.Therefore, treatments that dampen but do not eliminate the function of MDA-5 will likely elicit both an anti-CBV response in a manner that protects susceptible hosts from viral infection and yet do not allow the development of T1D.
Preventative measures for individuals at risk of T1D are not available and current management of the disease is through repetitive insulin injections to maintain near-normal blood glucose levels. Unfortunately, poor glycemic control can still occur and cause severe complications including blindness, heart disease, kidney and nerve problems. Researchers have made great progress toward the discovery of a preventative therapeutic for T1D, especially in the case of identifying potential causative factors driving the disease, though an exact etiology or promising preventative measure for the disease remains unclear. The recently identified common nsSNP and some rare variants in the IFIH1 gene responsible for producing the protein MDA-5 and likely protecting otherwise susceptible people from T1D, is a promising step in the right direction for developing a T1D preventative treatment. Our lab’s functional studies silencing MDA-5 in a CBV-induced T1D model will help uncover not only a link between a human polymorphism and susceptibility to T1D, but further connect innate immunity viral sensing with disease susceptibility that will hopefully lead to a potential target for T1D therapy.
ON BECOMING AN OCTOGENARIA: MATTERS OF HISTORY by Julian Davies
It is an interesting aspect of microbiology that because it is a relatively young science even the youngest faculty member in M&I will have met some of the “greats” in the field. However, it seems to me that there is not much interest in the history of the field; even graduate students, who employ all types of kits based on early discoveries, don't seem to know what they are using and why. For example, what are the restriction enzymes used in cloning kits, where did they come from and what are their roles in the biology of the bacteria that make them. (actually, for ease of production they are all cloned into E. coli these days, but this host is the natural source of only a few).
The history of microbiology is filled with amazing characters and great stories. How did its various aspects commence and become developed? What were the landmark discoveries? Did they follow on from each other in a logical manner? Who were the main characters (good and bad)? Many microbiologists have been awarded a Nobel Prize for their studies with microbes: who won the prize and who deserved it?
As most of you may be aware, I am not a microbiologist by training, but by osmosis. My BSc and PhD were in chemistry. I did carry out work on the chemical synthesis and structure determination of some interesting fungal products but had little idea what I was doing when I was growing Aspergillus strains! Or where the fungus came from and what it did. Nonetheless as my career developed I drifted more and more towards the life sciences, primarily microbiology. The turning point came when I decided that I did not enjoy chemistry and left a position at the University of Manchester to become a postdoc in the Department of Bacteriology and Immunology at Harvard Medical School in Boston. Bacteriology was completely new to me. And immunology… well, I had no idea what that was!
Fortunately, Hugh McDevitt, now a well-known immunologist at Stanford, was at HMS; we often lunched together and also played the occasional game of golf. Since we spent a lot of time looking for lost balls, he was able to explain some of the mysteries of immunology to me. Bacteriology was becoming increasingly molecular in the early 1960s (to the chagrin of the purists) and E. coli was the easiest and favourite source of DNA, ribosomes, different RNA molecules and enzymes. The cell-free systems for studying the basic steps in protein synthesis and analyzing protein synthesis inhibitors were readily available and we made them all from scratch.
Learning microbiology/bacteriology was a different matter; I could not understand most of the departmental seminars I attended (even if I stayed awake!). Fortunately I was recruited to teach bacteriology to the first-year medical students and as long as I kept one lecture ahead of them things went well.
In addition, I had the chance to listen to and meet many of the great microbiologists of the time.The 1960s were exceptional years for the modernization of microbiology and the development of molecular biology. Everyone came to Boston and visited Harvard. I had the chance to meet Josh Lederberg, Salva Luria, Francis Crick, Sydney Brenner, Seymour Benzer, Wally Gilbert, Marshall Nirenberg, Gobind Khorana, Severo Ochoa, Bill Hayes, Ernst Chain (of penicillin fame), François Jacob and many others. I had no idea how famous these people were and I probably asked them dumb questions, but they graciously replied. It was a great time to experience and learn.
Jim Watson was interested in the work I was doing on ribosomes and was especially helpful to me. Elegant genetic studies at MRC Cambridge (UK) showed that the genetic code was a triplet code (three bases per amino acid) and molecular tools allowed the code to be deciphered in vitro. There was a race to identify coding triplets and new assignments came out weekly. The biological sciences were changing before our eyes! Much of this period, when the field was relatively small and molecular biology was not recognized as a discipline in its own right, is now forgotten. There were no 'molecular biology' departments. One notable biochemist stated that “molecular biology is doing biochemistry without a license”. Interestingly, the Department of Biochemistry at UBC was renamed Biochemistry and Molecular Biology in the early 1990s.
I moved to the University of Wisconsin in Madison in 1967 and started to focus my studies on antibiotic resistance mechanisms in bacteria. An early issue arose with a dispute over the reasons behind the extensive development of antibiotic resistance in hospitals and the relationship to antibiotic usage in animal husbandry. As you all must know, this argument is ongoing.
The 1970s were even more exciting than the 60s in many respects, in part due to political and ethical debate concerning science and its applications. The significant advance of the 70s was the development of recombinant DNA and the major controversy that followed this discovery. After Herb Boyer and Stan Cohen did the first 'cloning' experiment in 1973, we all started to use the same techniques. Together with many colleagues I had been working on plasmids and antibiotic resistance genes for some time, so the tools were available. It proved relatively easy to isolate crude preparations of restriction enzymes and we could do all the DNA cutting and splicing we needed. One of my colleagues in Madison decided to clone turkey DNA for US Thanksgiving!
Do our present students know all this: the furore over the “cloning and manipulating life” controversy and the ridiculous regulations? It may seem surprising when we all use the technology without a second thought these days, even in high schools. Do students understand the biology of those cloning kits? Do they need to? Although the controversy has died down somewhat, there is still a lot of fuss over genetically modified foods, especially in Europe. Are they natural or unnatural? Is this the reason why Boyer and Cohen have not received the Nobel prize? They are certainly deserving, but it is unlikely to happen now. The Nobel awards are not free from politics!
However, these and other matters should be discussed by scientists and not left to bioethicists,et al. This is especially true when we realize that one of the biggest problems we face is that of the overuse of biologicals such as antibiotics. The erosion of antibiotic therapy is ever-increasing and there seems to be little that can be done to reverse it. The end of the antibiotic era means the end of 'control' of infectious disease and it is being allowed to happen – like fighting in hockey.
Now we in the 21st century are witnessing a sea-change in the microbiological sciences. The metagenome-microbiome revolution is changing the history of microbiology rapidly and decisively; all that we learned before will be confined to archives: how will this change our science? Nothing from the past can match what is happening at this very time. The new microbiology is here. Enjoy it while you can!

 In the great cycle of (academic) life, our Department is always welcoming new students and faculty while others become part of our alumni family. This year we had two of our professors, George Spiegelman and Hung-Sia Teh ‘graduate’ to retirement, each with plans that will keep them extremely busy. At the same time, we welcomed two new faculty members, Martin Hirst and Georgia Perona-Wright. Martin spent the previous few years at the BC Genome Sciences Centre and in this post-genomic era is bringing epigenetics to our department. Georgia is originally from the UK and worked at the Trudeau Institute in the woods of upstate New York before joining us. She is an ‘integrative immunologist’ who is interested in determining how local versus systemic immune responses determine whether or not different kinds of infections are cleared. She brought with her to Vancouver many interesting infectious disease models as well as her own canoe. We are currently in the process of hiring two more faculty members in exciting new, cross-disciplinary areas, geomicrobiology and human microbial ecology. The geomicrobiologist is a joint hire with the Department of Earth & Ocean Sciences. They will bring new dimensions to our environmental microbiology research and will further develop our combined undergraduate program in oceanography and microbiology. Microbial ecology originated as an aspect of environmental microbiology and it is now realized that the approaches developed to study microbial communities in the oceans or soils can be applied to the microbial communities of our gut, lung, skin, etc. These microbial communities and their interactions with the host are key determinants of health and disease. The combined efforts of microbiologists, immunologists, genome scientists, and bioinformaticians will be required to fully understand these complex interactions.
In the great cycle of (academic) life, our Department is always welcoming new students and faculty while others become part of our alumni family. This year we had two of our professors, George Spiegelman and Hung-Sia Teh ‘graduate’ to retirement, each with plans that will keep them extremely busy. At the same time, we welcomed two new faculty members, Martin Hirst and Georgia Perona-Wright. Martin spent the previous few years at the BC Genome Sciences Centre and in this post-genomic era is bringing epigenetics to our department. Georgia is originally from the UK and worked at the Trudeau Institute in the woods of upstate New York before joining us. She is an ‘integrative immunologist’ who is interested in determining how local versus systemic immune responses determine whether or not different kinds of infections are cleared. She brought with her to Vancouver many interesting infectious disease models as well as her own canoe. We are currently in the process of hiring two more faculty members in exciting new, cross-disciplinary areas, geomicrobiology and human microbial ecology. The geomicrobiologist is a joint hire with the Department of Earth & Ocean Sciences. They will bring new dimensions to our environmental microbiology research and will further develop our combined undergraduate program in oceanography and microbiology. Microbial ecology originated as an aspect of environmental microbiology and it is now realized that the approaches developed to study microbial communities in the oceans or soils can be applied to the microbial communities of our gut, lung, skin, etc. These microbial communities and their interactions with the host are key determinants of health and disease. The combined efforts of microbiologists, immunologists, genome scientists, and bioinformaticians will be required to fully understand these complex interactions.  If you ever see me driving “no hands”, just get out of the way - I earned my license in Istanbul eons ago! These days, in a contemplative mood upon my retirement, I often drive down memory lane wondering how I got half way around the world to Vancouver:
If you ever see me driving “no hands”, just get out of the way - I earned my license in Istanbul eons ago! These days, in a contemplative mood upon my retirement, I often drive down memory lane wondering how I got half way around the world to Vancouver: The Microbiology Immunology Student Association, otherwise known as MISA, carries 27 years of traditions and responsibilities. Each year, we uphold the decade-old goal: promote a fun, academic and social atmosphere for microbiology enthusiasts. While the scope and number of our activities have changed over the years, the basis remains. We cater to the students by hosting a handful of social events throughout the year, and provide guidance through their journey here at UBC.
The Microbiology Immunology Student Association, otherwise known as MISA, carries 27 years of traditions and responsibilities. Each year, we uphold the decade-old goal: promote a fun, academic and social atmosphere for microbiology enthusiasts. While the scope and number of our activities have changed over the years, the basis remains. We cater to the students by hosting a handful of social events throughout the year, and provide guidance through their journey here at UBC.

 It is an interesting aspect of microbiology that because it is a relatively young science even the youngest faculty member in M&I will have met some of the “greats” in the field. However, it seems to me that there is not much interest in the history of the field; even graduate students, who employ all types of kits based on early discoveries, don't seem to know what they are using and why. For example, what are the restriction enzymes used in cloning kits, where did they come from and what are their roles in the biology of the bacteria that make them. (actually, for ease of production they are all cloned into E. coli these days, but this host is the natural source of only a few).
It is an interesting aspect of microbiology that because it is a relatively young science even the youngest faculty member in M&I will have met some of the “greats” in the field. However, it seems to me that there is not much interest in the history of the field; even graduate students, who employ all types of kits based on early discoveries, don't seem to know what they are using and why. For example, what are the restriction enzymes used in cloning kits, where did they come from and what are their roles in the biology of the bacteria that make them. (actually, for ease of production they are all cloned into E. coli these days, but this host is the natural source of only a few).